Teknisk Analyse
Bioreaktivitet fra radiofrekvente strålingskilder i non-termiske intensiteter har i årtier været et særdeles omdiskuteret emne. Inden for de senere år er der dog frembragt megen ny viden som giver et - i hvert fald rudimentært - overblik over de væsentligste påvirkningsmekanismer. Meget af den nyere forskning er desværre endnu ikke blevet udmøntet i reviderede eksponeringsbegrænsende retningsliner, hvilket er baggrunden for Elektrobiologisk Selskabs fokus på anvendelse af forsigtighesprincippet i forhold til den almene befolknings eksponering.
Fotonenergi
I klassisk fysik og biologi har man traditionelt lagt megen vægt på fotonens energiniveau i forhold til at kategorisere mulig bioreaktivitet og deraf følgende skadevirkninger fra elektromagnetisk stråling.
Fotonenergien er særdeles enkel at udregne, idet Ef er fotonenergien, h er Planck's konstant og ν er frekvensen.
Ef=h⋅ν
Når fotonenergien er under 1eV har fotonen under normale omstændigheder ikke energi nok til at bryde molekylebindinger direkte. Strålingen kaldes i så tilfælde non-ioniserende. Traditionelt har man kun opereret med muligheden for termiske skadevirkninger ved non-ioniserende stråling. Inden for de senere år er der dog påbegyndt et decideret paradigmeskifte, hvor man i dag har en begyndende forståelse af nogle af de ekstremt komplicerede mekanismer, hvormed koherente påvirkninger langt under ioniseringsenergien kan medføre betydelig påvirkning af biologiske systemer.
En af de væsentligste og mest veldokumenterede påvirkningsmekanismer er det elektriske felts påvirkning af ionkanaler i cellemembraner. I cellemembranen findes forskellige proteinkomplekser, som medierer ioners adgang gennem cellemembranen. I denne forbindelse er der specielt fokus på calcium-ionen og de dertil hørende ionkanaler, kaldet VGCC (Voltage Gated Calcium Channels)
Exogene elektriske felters aktivering af VGCC'ere
I det følgende præsenteres en betydelig forenklet beregning af påvirkningsprincippet for interaktionsmekanismen mellem non-ioniserende radiofrekvent stråling og cellemembranens ionkanaler. For en fuld behandling af emnet henvises til originallitteratur[1], hvilken samtidigt er grundreference for nedenstående beregningsstruktur.
Følgende konstanter og udtryk anvendes i beregningen:
E=E0⋅sin ωt : Det elektriske felt (V/m)
x : Ion forskydning ved påvirkning af et elektrisk felt (m)
z : Valens
qe : Elementarladningen
λ : Dæmpningsfaktoren for ionbevægelsen i det pågældende medium
A : Amplitude i ionvibrationen (m)
F : Kraft, kraftpåvrikning (N)
ε0 : Dielektrisk konstant for vakuum
ε : Relative dielektiske konstant
∂∆Ψ : Krævet transmembranspænding for aktivering af VGCC
s : Membrantykkelse (m)
q : Ladningen på S4 spændingssensor
r : Afstanden mellem den beregnede frie ion med ladningen zqe og ladningen q på S4 spændingssensor
∂r : Mindste krævede ionbevægelse for aktivering af VGCC
ν : Frekvens (Hz)
Det kan vises[1], at en ions spatiale forskydning ved udsættelse for et elektrisk felt kan beskrives ved følgende udtryk:
x = ( E0zqe / λω ) cos ωt - ( E0zqe / λω )
Idet sidste del af udtrykket er konstant, vil ionens vibrationsbevægelse derfor kunne beskrives ved:
x = ( E0zqe / λω ) cos ωt
Det ses, at ionbevægelsen er faselåst til det eksterne felt, hvorfor amplituden i ionbevægelsen samtidigt kan beskrives ved udtrykket:
A = E0zqe / λω
Det ses her, at amplituden er proportionel til intensiteten og omvendt proportionel til frekvensen af det elektriske felt.
Det er samtidigt vist[2], at kraftpåvirkningen på en S4 spændingsensor i en VGCC kan beskrives ved følgende udtryk:
∂F = ∂∆Ψ ⋅ q/s
Idet ∂∆Ψ = 30mV[2] og q ≅ 1,7 qe [1] bliver kraftpåvirkningen, ∂F, for at åbne en enkelt VGCC 8,16×10-13 N.
Kraftpåvirkningen på en S4 fra en enkelt vibrerende ion kan herefter beregnes:
F = 1 / ( 4πεε0 ) ⋅ ( q⋅zqe ) / r²
Det ses samtidigt, at den mindste krævede ionbevægelse for at aktivere VGCC'en kan skrives ved:
∂r = 2πεε0∂F⋅r3 / q⋅zqe
Idet ε0 kan variere mellem 4 og 80[2; 3] bliver den mindste krævede ionbevægelse for at aktivere VGCC'en at regne i få picometer; 0,8×10-10 m for ε = 80 og 4×10-12m for ε = 4.
Ethvert eksternt elektrisk felt, som kan frembringe en ionvibration med en amplitude over 4x10-12m, vil derfor kunne aktivere VGCC'ere med deraf følgende elektrokemisk forstyrrelse af cellen, hvilket samtidigt betyder, at et elektrisk felt som overholder følgende beregning, vil have bioreaktivt potentiale i forhold til elektrokemisk forstyrrelse af cellen:
E0qe / λω > 4⋅10-12m
Herved kan det samtidigt ses, at et elektrisk felt med en intensitet over
Ε0 ≥ νx10-3
vil medføre bioreaktivt potentiale for en enkelt ion, hvorimod et pulseret felt vil kunne medføre bioreaktivt potentiale for en Ca2+ allerede ved
Ε0 ≥ ν⋅0,625⋅10-4
Frekvensen har herved afgørende betydning for feltets bioreaktivitet, idet det dog erindres, at moderne mikrobølgebaseret telekommunikation benytter stærk pulsmodulation, hvorfor den relevante frekvens for beregning af bioreaktivt potentiale ikke er bærebøglefrekvensen, men derimod de ELF subkomponenter, der kan udmåles i signalet.
Udregning af ELF-subkomponentfrekvenser i digitalt pulsmodulerede signaler
Digital pulsmodulation skaber forskellige lavfrekvente mønstre i det elektromagnetiske signal. Den digitalt indlejrede information i signalstrukturen kan kun afkodes med en avanceret digitalmodtager, men selve pulsstrukturen kan let afkodes med simple analoge instrumenter - og kan derved forårsage biologiske påvirkninger, idet vi, blandt andet via ovenstående udregning, har set, hvorledes der ved påvirkning af VGCC'ere i princippet foretages en analog afkodning af de lavfrekvente mønstre (kaldet ELF subkomponentstrukturer) i signalet.
Den velkendte GSM modulationsstruktur kan anvendes som indledende eksempel for at konkretisere problemfeltet. GSM er bygget op omkring en kombination af to forskellige måder at organisere radiotransmissioner på. Den ene af disse måder kaldes FDMA (frequency division multiple access), og den anden kaldes TDMA (time division multiple access). Kombinationen af FDMA og TDMA er valgt for at give plads til så mange samtaler som muligt på så få radiofrekvenser som muligt, idet operatørene skal betale staten for adgang til frekvensbåndene.
FDMA-strukturen giver således mulighed for omkring hundrede samtidige kommunikationsbånd inden for det allokerede frekvensbånd. Der er omkring 200kHz mellemrum mellem de forskellige kommunikationsbånd. For at give plads til endnu flere samtaler benyttes TDMA-strukturen oven på FDMA. TDMA bryder hvert kommunikationsbånd (GSM "multiframe" på 120 millisekunder) op i 8 tidsenheder, således at 8 mobiltelefoner samtidigt kan bruge samme kommunikationsbånd.
En konsekvens heraf er, at den enkelte mobiltelefon opererer med en transmissionscyklus på 1:8, således at den kun sender i en ottendedel af tiden, den er online. Samtidigt vil en basestation i en mobilmast (ud over normalt at sende med højere effekt) også have en markant højere (8/8) transmissionscyklus end en mobiltelefon. Basestationen kan altså forventes at sende hele tiden.
Disse pulseringsformer giver to distinkte ELF (Extremely Low Frequency) -komponenter i uplink GSM-signalet, nemlig en på 8,33Hz, idet hvert 26. signalpuls udelades og en på 217Hz, idet telefonen sender hvert 4,6ms med varighed af 0,57ms.
Signalerne og deres pulseringsform kan iagttages på nedenstående grafiske sammenstilling
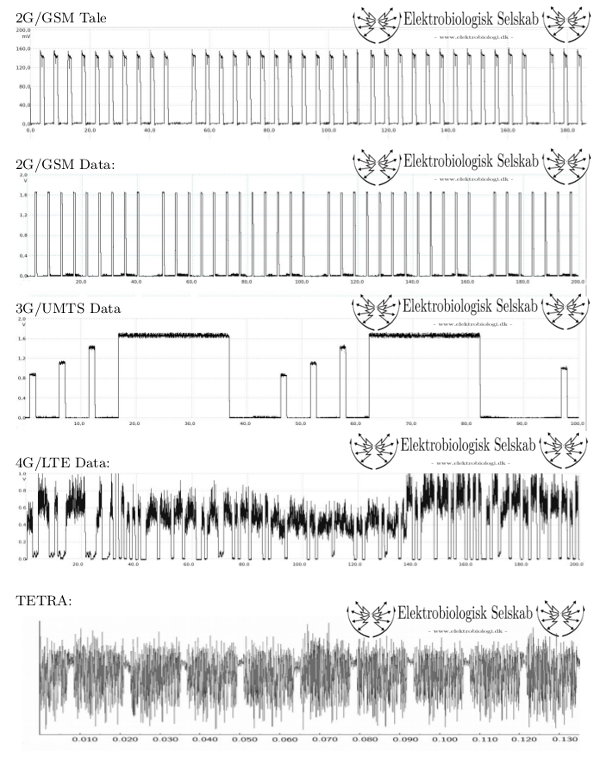
Ved indsættelse af ELF-subkomponentfrekvenserne for GSM i slutligningen
Ε0 ≥ ν⋅0,625⋅10-4
fåes således en nedre værdi for elektrisk feltintensitet for potentiel bioreaktivitet på ca 0,0005 V/m ved en ELF-subkomponentfrekvens på 8,33Hz og en tilsvarende nedre værdi for elektrisk feltintensitet for potentiel bioreaktivitet på ca 0,014 V/m ved en ELF-subkomponentfrekvens på 217Hz.
Kun signalstrukturer baseret på TDMA har faste mønstre for ELF-subkomponentfrekvenser. UMTS (3G) og LTE (4G) er ikke TDMA-baserede, hvorfor de præcise ELF-subkomponenter varierer afhængigt af den transmitterede datamængde. Ved praktisk måling varierer ELF-subkomponenter for disse signaler mellem nogle få hertz og nogle få kilohertz. Som det er tydeligt fra ovenstående beregning, er det klart de lavere ELF-subkomponentfrekvenser, der har den højeste bioreaktivtet.
En anden meget anvendt signalteknologi baseret på TDMA findes i den danske beredskabsradio TETRA (som i DK kaldes SINE). Her findes ELF-subkomponenter på 16 Hz og 70 Hz, hvilket igen svarer til nedre elektrisk feltintensiterer for potentiel bioreaktivitet på 0,001 V/m og 0,004 V/m.
Som betragtningsbaggrund for ovenstående beregninger kan det nævnes, at den officielle eksponeringsgrænse for non-ioniserende radiofrekvent stråling i frekvensområdet fra 2-300GHz er 61 V/m[4], hvilket samtidigt implicerer at den politisk vedtagne grænseværdi er 122000 gange højere end den beregnede nedre grænse for potentiel bioreaktivitet.
Den ganske betydelige forskel, mellem den politisk vedtagne grænseværdi og den forskningsmæssigt begrundede nedre grænse for potentiel bioreaktivitet, er hovedbaggrunden for Selskabets fastholdelse af vigtigheden af anvendelse af forsigtighedsprincippet i forhold til den almene befolknings eksponering
Referencer
[1] Panagopoulos, DJ. Biological impacts, action mechanisms, dosimetry and protection issues of mobile telephony radiation. In Barnes MC and Meyers NP, eds. Mobile Phones: Technology, Networks and User Issues. New York: Nova Science Publishers, 2011
[2] Panagopoulos DJ, Messini N, Karabarbounis A, Filippetis AL, and Margaritis LH. 2000. A Mechanism for Action of Oscillating Electric Fields on Cells. Biochemical and Biophysical Research Communications, 272(3), 634-640.
[3] Honig BH, Hubbell WL, Flewelling RF. 1986. Electrostatic Interactions in Membranes and Proteins. Ann.Rev.Biophys.Biophys.Chem.,15.
[4] ICNIRP. 1998. Guidelines for limiting exposure to time-varying electric, magnetic, and electromagnetic fields (up to 300 GHz). International Commission on Non-Ionising Radiation Protection. Health Physics 74, 494‐522.